Biochimica Ferorelli
Nei mammiferi l’irrorazione ematica dei tessuti è garantita da una estesa rete di vasi, collegata al cuore, costituita da arterie, arteriole, capillari, venule, vene e sistema linfatico, che nel loro insieme definiscono l’apparato cardiovascolare. Anomalie a livello vascolare intervengono nella storia naturale delle più svariate patologie, rappresentando il “fattore di terreno” che ne favorisce lo sviluppo: ciò consente l’identificazione di un approccio unitario mirato al riequilibrio e definisce il campo di ricerca della nuova Enzimologia Clinica Biodinamica in Patologia generale.
Introduzione: aspetti emodinamici
I capillari rappresentano il principale sito di scambio, per diffusione, fra sangue e tessuti; la maggior parte delle cellule si trova a una distanza di poche decine di micron dai capillari, questi vasi formano una rete vascolare più sviluppata e a più intimo contatto tissutale, e la loro attività è direttamente proporzionale a quella dei tessuti con cui sono collegati. In alcuni tessuti, peraltro, i capillari non penetrano direttamente e i gradienti di diffusione possono avere una lunghezza di centinaia di micron, come nelle cartilagini, in alcuni epiteli, nel tessuto connettivo della cornea e della dentina. Prima di entrare nella descrizione riguardante l’organizzazione dei canali vascolari, è opportuno richiamare l’attenzione sulla biologia dei componenti cellulari circolanti nel sistema sanguifero e linfatico nonché il loro sviluppo nel midollo osseo e nel tessuto linfatico. La componente cellulare del sangue è costituita da cellule della serie rossa (eritrociti), di origine midollare ossea, che veicolano l’ossigeno e, in misura minore, l’anidride carbonica; cellule della serie bianca (leucociti), rappresentate da una grande varietà di elementi cellulari con funzioni difensive, e da cellule della serie piastrinica che intervengono nell’emostasi. La componente linfoide comprende cellule che vengono formate sia nel midollo osseo che in molti altri siti al di fuori di esso, quali il timo, i linfonodi, la milza e il tessuto linfoide associato a tratti del tubo gastroenterico e dei bronchi (follicoli linfatici). Le pareti del sistema vascolare sono tappezzate da cellule endoteliali (che saranno più avanti descritte), per la loro normale attività queste cellule necessitano di un continuo equilibrio fra l’introduzione di sostanze nutritizie e l’emissione dei prodotti cellulari e dei cataboliti. Nei vertebrati, uomo compreso, il liquido dell’organismo è suddiviso mediante confini semipermeabili in quattro compartimenti: plasma sanguigno, linfa, liquido interstiziale e liquido intracellulare; le grosse molecole sono per lo più trattenute nel sangue, l’acqua e le sostanze in essa disciolte, circolano negli spazi extravascolari che si trovano fra le cellule dei tessuti e vengono a costituire il liquido tissutale. I vasi che portano il sangue verso i polmoni o ritornano da questi costituiscono la circolazione polmonare, mentre quelli che portano il sangue o lo raccolgono dal resto dell’organismo formano la circolazione generale o sistemica. In entrambe le circolazioni il sangue viene pompato e fatto progredire nelle arterie, che mediante ramificazioni successive aumentano di numero e diminuiscono di calibro fino a diventare arteriole, che si risolvano in una rete di capillari; a livelli di questi fini vasi avvengono i principali scambi fra plasma sanguigno e liquido interstiziale, il sangue ritorna la cuore attraverso vene e venule confluenti. Il sistema circolatorio linfatico trasporta la linfa dagli intersizi tissutali fino alle vene situate alla base del collo; questa circolazione drena il liquido interstiziale e gli elementi cellulari mobili in esso contenuto nei capillari linfatici che terminano a fondo cieco, essi convergono in vasi linfatici di vario calibro i quali portano la linfa al sistema sanguigno venoso. Assicurando la distribuzione dei metaboliti a tutti i tessuti e a tutte le cellule, il sistema circolatorio contribuisce, assieme al sistema nervoso, al collegamento ed all’integrazione di tutti i costituenti dell’organismo.
Caratteristiche del sangue
Il sangue è un tessuto connettivo liquido eterogeneo, composto da una parte fluida chiara (plasma)e da una parte corpuscolata (corpuscoli): a causa di questa mescolanza si comporta, da un punto di vista idrodinamico, in modo complesso e appartiene a quella classe di fluidi che vengono definiti non Newtoniani. Il plasmaè un liquido chiaro e lievemente giallastro che contiene molte sostanze in soluzione o in sospensione; è ricco di ioni sodio e cloro e contiene inoltre potassio, calcio, magnesio, fosfati, bicarbonati e altri ioni, glucosio, amminoacidi, ecc. I colloidi comprendono proteine plasmatiche ad lato peso molecolare, protrombina, immunoglobuline, proteine coinvolte nella difesa immunitaria, ecc., che saranno più avanti descritte. Gli elementi del sangue sono riuniti in tre gruppi: gli elementi della serie rossa, della serie bianca e le piastrine.
Elementi cellulari della serie rossa: Eritrociti.
Rappresentano la maggior parte degli elementi corpuscolari del sangue (il 99% del numero totale); ogni cellula ha la forma di un disco biconcavo, sono prive di nucleo e mostrano una tendenza ad aderire reciprocamente ai bordi e a formare delle pile fluttuanti di cellule.
Membrane biologiche e trasporto degli eritrociti.
Le membrane e i lipidi polari rappresentano la quasi totalità della massa delle membrane biologiche; la piccola quantità dei carboidrati presenti nelle membrane fanno parte in genere delle glicoproteine o dei glicolipidi. La quantità relative dei lipidi e di proteine variano a seconda del tipo di membrana e riflettono la differenza delle loro funzioni biologiche; i rivestimenti di mielina che servono come isolanti elettrici passivi avvolti intorno a certi neuroni, sono sostituiti principalmente da lipidi, mentre le membrane dei batteri, dei mitocondri e dei cloroplasti, in cui hanno luogo molti processi metabolici catalizzati da enzimi, contengono più proteine che lipidi. Tutte le membrane biologiche presentano alcune proprietà fondamentali: sono impermeabili alla maggior parte dei soluti carichi o polari, ma sono permeabili ai composti non polari; hanno uno spessore che varia da 5 a 8 nm; appaiono composte da tre strati quando sono viste in sezione trasversale al microscopio elettronico. Sulla base delle prove sperimentali, sulla composizione chimica e sugli studi fisici sulla permeabilità e sugli spostamenti delle singole molecole, si prospetta la formulazione del modello a mosaico fluido: vedi figura sotto riportata.
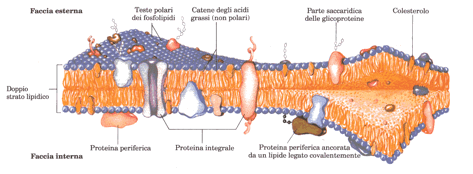
Modello del mosaico fluido per la struttura della membrana.
Le catene degli acidi grassi all’interno della membrana formano una regione idrofobica fluida; le proteine integrali galleggiano in questo mare di lipidi mediante interazioni idrofobiche tra le catene laterali dei loro residui amminoacidici non polari e le catene degli acidi grassi. Sia le proteine che i lipidi sono liberi di spostarsi lateralmente nel piano del doppio strato, mentre i movimenti da una faccia all’altra della membrana sono meno frequenti. I gruppi saccaridici legati ad alcuni lipidi e proteine della membrana plasmatica sono sempre esposti sulla faccia extracellulare della membrana.
I lipidi di membrana sono distribuiti nelle due facce del doppio strato in modo asimmetrico, anche se, a differenza delle proteine, l’asimmerreia non è assoluta; nelle membrane plasmatiche, ad es., certi tipi di lipidi si trovano principalmente nella faccia esterna del doppio strato, mentre altri sono nella faccia interna (citoplasmatica).
Anche se la struttura a doppio strato è di per sé stabile, le singole molecole di fosfolipidi e di steroli hanno una grande libertà di movimento all’interno del piano della membrana: vedi figura.
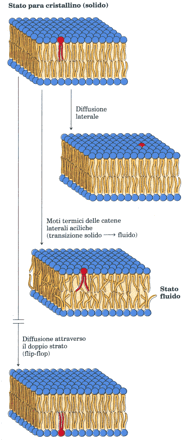 esse hanno movimenti di diffusione laterale così veloci che una molecola di lioide può circumnavigare un eritrocita in pochi secondi.
esse hanno movimenti di diffusione laterale così veloci che una molecola di lioide può circumnavigare un eritrocita in pochi secondi.
Il grado di fluidità dipende dalla composizione lipidica e dalla temperatura; a bassa temperatura i movimenti sono molto limitati e il doppio strato è una struttura quasi cristallina (paracristallina).
Sopra una certa temperatura che è caratteristica per ogni tipo di membrana, i lipidi possono muoversi liberamente e velocemente; la temperatura di transizione da uno stato paracristallino solido a quello fluido dipende dalla composizione lipidica della membrana.
Gli acidi grassi saturi si impacchettano bene nelle strutture paracristalline, mentre i ripiegamenti nelle catene di acidi grassi insaturi interferiscono con l’impacchettamento e impediscono la formazione di uno strato paracristallino: più elevata è la proporzione di acidi grassi saturi e più elevata è la temperatura di transizione solido-fluido della membrana.
Il contenuto in steroli è un altro determinante della temperatura di transizione di una membrana; la struttura rigida e planare del nucleo steroideo inserita fra la catena degli acidi grassi produce due effetti sulla fluidità: sotto la temperatura di transizione solido-fluido l’inserzione dello sterolo impedisce una disposizione altamente ordinata della catene degli acidi grassi e quindi fluidifica le membrane.
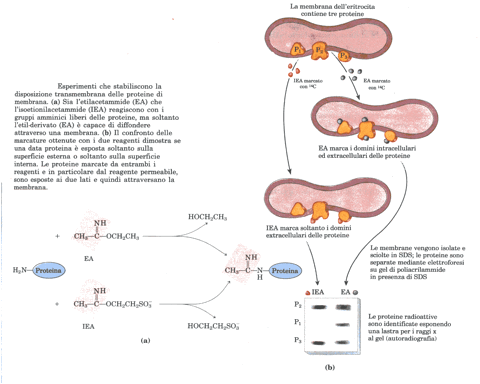
A una temperatura superiore s quella di transizione, il sistema ad anelli rigidi dello sterolo riduce la libertà di movimento delle catene di acido grasso vicine intorno ai legami carbonio-carbonio, e riduce al tempo stesso la fluidità del nucleo interno del doppio strato: gli steroli servono quindi a moderare gli estremi, cioè la solidità e la fluidità assolute delle membrane che li contengono. Mentre i movimenti laterali o le flessioni termiche delle catene acicliche possono avvenire liberamente, un terzo tipo di movimento presenta invece notevoli restrizioni: la diffusione attraverso il doppio strato o flip-flop, il movimento di un lipide da una faccia della membrana all’altra, un processo che determina una grande variazione positiva di energia positiva. Vi sono proteine che facilitano la diffusione a flip-flop, generando una via di trasferimento transmembrana energeticamente più favorevole; sotto questo aspetto l’eritrocita umano è particolarmente conveniente in quanto l’unica membrana presente in queste cellule è quella plasmatica. Esperimenti simili a quelli descritti nella figura sotto riportata hanno dimostrato che la molecola di glicoforina attraversa la membrana dell’eritrocita; il suo dominio amminoterminale (che contiene i carboidrati) è sulla superficie esterna dell’eritrocita, mentre il suo dominio carbossiterminale si estende all’interno della cellula.
I domini carbossiterminali e amminoterminale contengono molti residui amminoacidici polari o carchi e sono quindi relativamente idrofilici; un lungo segmento al centro della molecola della glicoforina contiene soltanto però residui amminoacidi idrofobici, da queste osservazioni sperimentali è derivata l’organizzazione molecolare della glicoforina (V. figura) – La disposizione della glicoforina nella membrana dell’eritrocita.
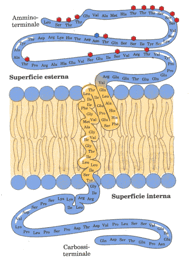 Un dominio idrofilico, contenete tutti i residui saccaridici, è sulla superficie esterna e un altro domino idrofilico è sulla superficie interna. Gli esagoni rappresentano unità tetrasaccaridiche (contenenti due NeuNAc, Gal e GalNAc) legate con un legame O-glicosidici a residui di serina o di treonina della proteina; l’esagono in blu rappresenta un oligosaccaride legato con un legame N-glicosidico a un residuo di asparagina. Un segmento composto da residui relativamente idrofobici forma una α elica che attraversa il doppio strato lipidico.
Un dominio idrofilico, contenete tutti i residui saccaridici, è sulla superficie esterna e un altro domino idrofilico è sulla superficie interna. Gli esagoni rappresentano unità tetrasaccaridiche (contenenti due NeuNAc, Gal e GalNAc) legate con un legame O-glicosidici a residui di serina o di treonina della proteina; l’esagono in blu rappresenta un oligosaccaride legato con un legame N-glicosidico a un residuo di asparagina. Un segmento composto da residui relativamente idrofobici forma una α elica che attraversa il doppio strato lipidico.
Dai risultati ottenuti dagli studi sulla glicoforina è possibile trarre un’altra conclusione: essa non si sposta da una faccia all’altra del doppio strato lipidico; la sua disposizione nella membrana è asimmetrica ne consegue che un orientamento mediante una diffusione a flip-flop accade solo molto di rado per non dire mai. Le proteine di membrana possono essere divise in due gruppi: proteine integrali, saldamente legate alla membrana, proteine periferiche, legate alla membrana solo debolmente. Le proteine integrali di membrana hanno in genere domini ricchi di amminoacidi idrofobici; in alcune proteine vi è una singola sequenza idrofobica nel centro della proteina (come nella glicoforina) oppure nella regione ammino-o-carbossi-terminale; altre proteine di membrana hanno diverse sequenze idrofobiche, ognuna sufficientemente lunga da attraversare il doppio strato lipidico quando sono nella conformazione ad α elica. Molte proteine sono tenute associate alla membrana da interazioni elettrostatiche e da legami idrogeno con i domini idrofolici delle proteine integrali di membrana ; le proteine periferiche possono servire come regolatori degli enzimi legati alla membrana, come ponti di connessione tra le proteine integrali di membrana e le strutture intracellulari oppure come limiti alla mobilità di certe proteine di membrana. Ogni cellula deve recuperare dal suo ambiente circostante il materiale grezzo che le serve per la biosintesi e per la produzione di energia e deve rilasciare sempre nell’ambiente gli scarti del metabolismo; la membrana plasmatica contiene proteine che riconoscono in modo specifico e trasportano all’interno della cellula sostanze come gli zuccheri, gli amminoacidi e gli ioni inorganici. A parte poche eccezioni, il traffico delle piccole molecole attraverso la membrana plasmatica avviene ad opera di processi mediati da proteine, mediante canali transmembrana, trasportatori o pompe. Il passaggio transmembrana di composti polari e di ioni viene reso possibile da proteine di membrana che abbassano l’energia di attivazione per il trasporto, generando una via alternativa di attraversamento per uno specifico soluto; le proteine che hanno la capacità di produrre questa diffusione facilitata o trasporto passivo non sono enzimi nel senso comune della parola; esse spostano i loro substrati da un compartimento all’altro, ma non li modificano chimicamente, queste proteine sono chiamate trasportatori o permeasi.
Il metabolismo che produce energia nell’eritrocita dipende da un rifornimento costante di glucosio dal plasma sanguigno, dove la concentrazione dello zucchero viene mantenuta intorno a 5 mM; il glucosio entra nell’eritrocita per diffusione facilitata attraverso una specifica glucoso-permeasi. Questa proteina integrale di membrana ha 12 segmenti idrofobici e probabilmente attraversa la membrana 12 volte; consente al glucosio di entrare nella cellula ad una velocità circa 50.000 volte maggiore di quella della diffusione non assistita attraverso il doppio strato assistito. L’eritrocita contiene un altro sistema di diffusione facilitata, uno scambiatore di anioni, che essenzialmente è il trasportatore della CO2 prodotta dai tessuti in attività come il muscolo, il fegato e i polmoni; la CO2 prodotta viene rilasciata nel plasma sanguigno ed entra nell’eritrocita, dove viene convertita in bicarbonato (HCO3-) ad opera dell’enzima anidrasi carbonica, il bicarbonato passa a sua volta nel plasma per essere trasportato ai polmoni. Lo scambiatore cloruro-bicarbonato, chiamato anche proteina scambiatrice di anioni, aumenta la permeabilità della membrana dell’eritrocita all’HCO3- di un fattore pari a circa un milione di volte; come il trasportatore del glucosio, è una proteina integrale di membrana che attraversa probabilmente la membrana 12 volte, ma a differenza del trasportatore di glucosio, questa proteina produce un trasporto bidirezionale: per ogni ione bicarbonato che si muove in una direzione, uno ione Cl- si sposta nella direzione opposta. Vedi figura sotto riportata.
 Lo scambiatore cloruro-bicarbonato della membrana dell’eritrocita consente l’ingresso o l’uscita della HCO-3 senza variazioni nel potenziale elettrico transmembrana.
Lo scambiatore cloruro-bicarbonato della membrana dell’eritrocita consente l’ingresso o l’uscita della HCO-3 senza variazioni nel potenziale elettrico transmembrana.
La funzione di questo sistema di trasporto è quello di aumentare la capacità di trasporto della CO2 nel sangue.
Quando, come in questo caso, i due soluti si muovono in condizioni opposte, il processo viene detto antiporto; nel sinporto, i due substrati si muovono contemporaneamente nella stessa direzione
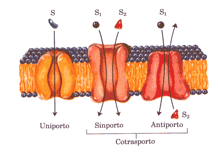 Le tre classi di sistema di trasporto differiscono nel numero di soluti (substrati) trasportati e nella direzione in cui ogni substrato viene trasportato; questa classificazione non tiene conto della eventuale richiesta di energia del sistema (trasporto attivo) o della su dipendenza energetica (trasporto passivo.
Le tre classi di sistema di trasporto differiscono nel numero di soluti (substrati) trasportati e nella direzione in cui ogni substrato viene trasportato; questa classificazione non tiene conto della eventuale richiesta di energia del sistema (trasporto attivo) o della su dipendenza energetica (trasporto passivo.
Nel trasporto passivo la specie trasportata segue sempre la direzione del gradiente di concentrazione e non si ha accumulo oltre la concentrazione di equilibrio.
Il trasporto attivo, al contrario, accumula sempre il soluto su un lato della membrana; il trasporto attivo è termodinamicamente sfavorito (endoergonico) e ha luogo soltanto quanto è accoppiato a un processo esoergonico come l’assorbimento della luce, una reazione di ossidazione, la demolizione di ATP oppure il flusso concomitante di un’altra specie nella direzione del su gradiente elettrochimico. Nel trasporto attivo primario l’accumulo del soluto è accoppiato direttamente ad una reazione esoergonica, il trasporto attivo secondario avviene quando il trasporto endoergonico (controgradiente) è accoppiato a un flusso esoergonico (nella direzione del gradiente) di un soluto diverso che era stato in precedenza accumulato da un trasporto attivo primario su un lato della membrana. Tutte le cellule mantengono nel loro interno una concentrazione di Na+ più bassa e una concentrazione di K+più elevata delle concentrazioni di questi elementi nel mezzo circostante; questa differenza viene stabilita e poi mantenuta da un sistema primario di trasporto attivo presente nella membrana plasmatica e costituito dall’enzima Na+ K+ ATPasi, che accoppia la demolizione dell’ATP al contemporaneo movimento degli ioni NA+ e K+ contro i loro gradienti di concentrazione. Per ogni molecola di ATP convertita in ADP e Pi il trasportatore porta dentro la cellula due ioni K+ ed espelle 3 ioni Na+ , facendo ovviamente attraversare loro la membrana plasmatica; il meccanismo, anche se non ancora perfettamente conosciuto, è illustrato nella figura sotto riportata.
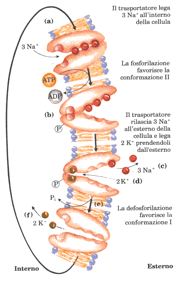 Il processo inizia con il legame di tre ioni Na+ a siti ad alta affinità sulla subunità più grande della proteina trasportatrice sulla superficie interna della membrana. (a) la stessa parte della subunità più grande possiede anche un sito che lega l’ATP.
Il processo inizia con il legame di tre ioni Na+ a siti ad alta affinità sulla subunità più grande della proteina trasportatrice sulla superficie interna della membrana. (a) la stessa parte della subunità più grande possiede anche un sito che lega l’ATP.
La fosforilazione del trasportatore modifica la sua conformazione (b) e diminuisce la sua affinità per Na+ , producendo il rilascio di questo ioni sulla superficie esterna della membrana (c).
In seguito gli ioni K+ all’esterno si legano ai siti ad alta affinità nella porzione extracellulare della subunità più grande (d); l’enzima viene quindi defosforilato, riducendo la sua affinità per il K+ (e) che viene rilasciato all’interno della cellula (f).
La proteina trasportatrice è di nuovo pronta per un altro ciclo di pompaggio degli ioni NA+ e K+.
Studiando le proteine di membrana dell’eritrocita umano mediante l’elettroforesi unidimensionale su gel di poliacrilamide SDS si rivelano circa 15 bande proteiche principali, con P.M. che variano fra 15.000 e 250.000. Tra di esse: la spettrina, la glicoforina e la banda III, rendono conto di oltre il 60% (in peso) della proteina di membrana totale. La maggior parte delle molecole proteiche della membrana degli eritrociti umani si trova esposta in un ambiente acquoso solamente sul lato interno (quello citoplasmatico) della membrana e, il più delle volte, basta una soluzione di bassissima forza ionica per distaccarle dalla membrana. La più abbondante fra le proteine di questo genere è la spettrina, lunga molecola fibrosa che costituisce circa il 30% della massa proteica della membrana (ogni cellula ne contiene approssimativamente 3×10 5 esemplari). La spettrina è costituita da un complesso di due grandissime catene polipeptidiche (ciascuna fra 200.000 e 240.000 dalton) disposte a mo di rete filamentosa sulla superficie citoplasmatica della membrana; esistono prove che alla costituzione di questa sorta di rete partecipano l’actina e almeno un’altra proteina e che la rete a maglia in questione contribuisca a mantenere la forma biconcava dell’eritrocita e a deformare le cellule quanto basta a consentire il passaggio lungo i capillari sottili. Tramite una proteina detta ankirina la spettrina interagisce indirettamente con l’estremo citoplasmatico delle proteine della banda III; ciò riduce di molto la velocità di diffusione di queste molecole della banda III nel piano della membrana. Vedi figura.
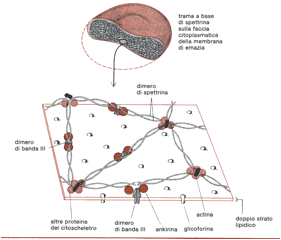 Le due catene polipeptidiche della spettrina sono disposte parallelamente e variamente avvolte l’una intorno all’altra a formare dimeri flessibili.
Le due catene polipeptidiche della spettrina sono disposte parallelamente e variamente avvolte l’una intorno all’altra a formare dimeri flessibili.
Sul versante citoplasmatico della membrana i dimeri della spettrina si congiungono testa a testa formando tetrameri saldati assieme in un reticolo di brevi segmenti di filamenti actinici e da altre proteine.
Attraverso un’altra proteina la ankirina, le molecole della spettrina interagiscono pure con le proteine della banda III.
Emoglobina.
 È una proteina globulare con un peso molecolare di 67.000, composta da molecole di globina legate all’eme, che contiene, a sua volta, un gruppo profirinico contenente ferro. Ogni molecola è costituita da quattro subunità: ognuna delle quali è a sua volta costituita da una catena polipeptidica avvolta a spirale con una fessura che contiene un singolo gruppo eme; nel sangue degli individui normali si possono trovare quattro catene polipeptidiche chiamate alfa, beta, gamma e delta. L’emoglobina A (HbA), che nell’adulto rappresenta la classe maggiore, contiene due catene alfa e due beta; l’emoglobina A2 (HbA2), presente in una minore percentuale di adulti, è formata da due catene alfa e due delta, l’emoglobina F (HbF) trovata ne feto e nei primi periodi di vita postnatale, consta di due catene alfa e due gamma. Nelle condizioni patologiche della thalassemia viene sintetizzato soltanto un tipo di catena, in modo tale che una molecola possa contenere quattro catene alfa (b-thalassemia) o, più comunemente, quattro catene beta (a-thalassemia).
È una proteina globulare con un peso molecolare di 67.000, composta da molecole di globina legate all’eme, che contiene, a sua volta, un gruppo profirinico contenente ferro. Ogni molecola è costituita da quattro subunità: ognuna delle quali è a sua volta costituita da una catena polipeptidica avvolta a spirale con una fessura che contiene un singolo gruppo eme; nel sangue degli individui normali si possono trovare quattro catene polipeptidiche chiamate alfa, beta, gamma e delta. L’emoglobina A (HbA), che nell’adulto rappresenta la classe maggiore, contiene due catene alfa e due beta; l’emoglobina A2 (HbA2), presente in una minore percentuale di adulti, è formata da due catene alfa e due delta, l’emoglobina F (HbF) trovata ne feto e nei primi periodi di vita postnatale, consta di due catene alfa e due gamma. Nelle condizioni patologiche della thalassemia viene sintetizzato soltanto un tipo di catena, in modo tale che una molecola possa contenere quattro catene alfa (b-thalassemia) o, più comunemente, quattro catene beta (a-thalassemia).
Nell’emoglobina a cellule falciformi, una sola alterazione della catena beta dell’emoglobina (la valina è sostituita dalla glutamina) determina una notevole modificazione nel comportamento degli eritrociti e della loro capacita di trasportare ossigeno.
Confronto fra la struttura di una globina a catena unica e quella di una globina a 4 catene; la globina a 4 catene rappresenta nella figura l’emoglobina, che è un complesso formato da due catene alfa e due beta.
Elementi cellulari della serie bianca.
I leucociti (globuli bianchi o cellule bianche) appartengono ad almeno 5 differenti categorie, differenziabili per dimensioni, forma del nucleo e inclusioni citoplasmatiche. I leucociti neutrofili polimorfonucleati (neutrofili, eterofili o polimorfi) rappresentano la maggior parte dei leucociti (60-70% negli adulti, con un numero di 3000-6000/ml). I neutrofili rappresentano un importante elemento del sistema di difesa dell’organismo; essi possono inglobare microbi e particelle presenti in circolo, e dopo aver migrato fra le cellule endoteliali che rivestono i capillari o le venule, da luogo a fagocitosi locale nei tessuti extravascolari, ovunque sia necessario.
L’inglobamento di elementi estranei è seguito dalla digestione, che avviene mediante la fusione di vacuoli fagocitari; essa è dovuta dapprima, a granuli specifici, una volta che, mediante un trasporto attivo di protoni, il pH è stato abbassato a 4, e successivamente, a granuli non specifici che portano a termine che portano a termine il processo di distruzione batterica e di digestione. Le reazioni che hanno luogo durante questi eventi comprendono: l’ossidazione e l’aggiunta di radicali alogenuri ai materiali inglobati, grazie all’azione di enzimi che hanno l’effetto di denaturare le proteine e le macromolecole; il lisozima e la lattoferina sono anche molto tossici per i batteri. In seguito alla fagocitosi, i granuli citoplasmatici dei neutrofili vanno incontro ad un graduale esaurimento e si produce una notevole riduzione del loro numero (degranulazione); i granuli possono essere anche emessi all’esterno della cellula, quando questa ultima viene stimolata in maniere appropriata, per danneggiare o uccidere organismi o cellule circostanti.
Leucociti eosinofili.
Queste cellule sono simili, per dimensione, forma e capacità di movimento, ai neutrofili, ma sono presenti solamente in piccole quantità nel sangue di soggetti normali (100-400/ml); i nuclei presentano due lobi prominenti collegati da un tratto prominente collegati da un tratto sottile; il citoplasma è gremito di granuli specifici, sferici o lievemente ellissoidali. La matrice contiene vari enzimi lisosomiali che comprendono la fosfatasi acida , la ribonucleasi, la fosfolipasi; sono in grado di passare dal circolo nei tessuti extravascolari, quando vengono stimolati in maniera opportuna. Aumentano sensibilmente in presenza di patologie allergiche.
Monociti.
Sono i leucociti agranulari più grandi ma rappresentano solo una piccola parte dei leucociti e sono presenti nel sangue in un numero di 100/770 /ml; il citoplasma forma un ampio orlo citoplsamatico attorno al nucleo; in vicinanza dell’incisura nucleare, si trova un vistoso complesso del Golgi e delle vescicole. Sono presenti molti lisosomi insieme al reticolo endoplsamtico rugoso periferico; i mitocondri sono assai abbondanti e riflettono la elevata attività motoria della cellula: i monociti sono cellule ad attività fagocitaria.
Linfociti.
Queste cellule sono il secondo tipo più numeroso di leucociti, rappresentando circa il 20-30% del totale; come gli altri leucociti, si trovano anche nei tessuti extravascolari, ma sono notevoli in quanto vengono ad essere formati in gran numero, sia all’esterno del midollo osseo che all’interno di esso. Vedi figura sotto riportata.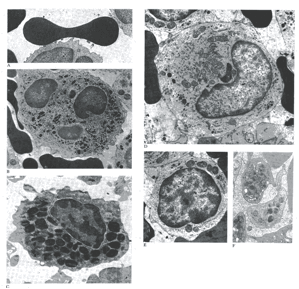
A): eritrocito che mostra il profilo biconcavo e il contenuto omogeneamente denso; B): neutrofilo che mostra il nucleo polilobulato che in sezione appare come profili nucleari separati; C): Eosinofilo, che mostra granulazioni specifiche con inclusioni cristalline; D): monocito che mostra una incisura nucleare, un reticolo endoplasmatico e dei lisosomi; E): piccolo linfocito; F): un gruppo di piastrine.
Piastrine.
Le piastrine del sangue, note anche come trombociti, sono dischi irregolari od ovali relativamente piccoli, presenti in gran numero nel sangue (250.000-300.000/ml).
Ogni piastrina è circondata da una membrana citoplasmatica con uno spesso rivestimento ricco di glicoproteine, responsabile della proprietà adesive delle piastrine; al di sotto della superficie vi è una banda di circa 10 microtuboli che girano attorno al perimetro della cellula e probabilmente ne determinano la forma. Associati a questi microtuboli ci sono i filamenti di actina, miosina e di altre proteine coinvolte nella contrazione cellulare; all’interno del citoplasma vi sono anche mitocondri, glicogeno e una certa quantità di reticolo endoplasmatico agranulare, compreso alcuni stretti canali tubulari e alcune invaginazione della membrana plasmatica e varie vescicole delimitate da membrana. I granuli alfa sono più cospicui e presentano diametri superiori a 500 nm; contengono un fattore di crescita di derivazione piastrinica (PGDF), fibrinogeno ed altre sostanze; nei granuli delta, molto più piccoli (330nm) c’è la 5-idrossitriptamina (serotonina) prelevata dal plasma e concentrata, ioni calcio, ADP, ATP e pirofosfato; nei granuli lambda, ancora più piccoli (250 nm) sono contenuti alcuni enzimi lisosomiali.
Le sostanze rilasciate dai granuli alfa, insieme ai fattori rilasciati dal tessuto danneggiato, danno il via a una complessa serie di reazioni chimiche nel plasma del sangue che porta alla formazione di filamenti insolubili di fibrina, che vengono a creare un reticolo tridimensionale chiamato coagulo piastrinico. Infatti, le piastrine aderiscono saldamente sui filamenti dell’intricata rete tridimensionale di fibrina e riempiono le maglie della rete, inserendo in profondità, negli spazi esistenti tra i filamenti, espansioni delle loro superfici (filopodi); successivamente le piastrine si contraggono (retrazione del coagulo piastrinico) per interazione dei filamenti di acrina con quelli di miosina, presenti all’interno del loro citoplasma, concentrando il coagulo di fibrina e riavvicinando fra di loro le pareti dei vasi sanguiferi su cui esse aderiscono. Alla fine, quando la parete del vaso è stata riparata, la rimozione del coagulo avviene come risultato di complesse attività svolte da parte di enzimi, come la plasmina, formata dagli attivatori del plasminogeno presenti nel plasma, e probabilmente coadiuvati da enzimi lisosomiali, derivati da piccoli granuli (granuli lambda) delle piastrine.
Il sistema linfoide.
I linfociti, nell’embrione, originano dapprima da cellule mesenchimali del sacco vitellino, e più tardi nel fegato e nella milza; queste primitive cellule staminali linfoidi, successivamente risiedono nel midollo osseo, che diventa il solo luogo in cui, dopo la nascita, proliferano le cellule staminali. Queste cellule si dividono in maniera differente per dare luogo a linfoblasti, che a loro volta si dividono per un certo numero di volte e si trasformano in piccoli linfociti; alcuni di questi passano nel sangue circolatorio e si portano verso il timo dove migrano e vanno a colonizzare i lobuli timici portando a termine il loro processo maturativo e vengono chiamati T-linfociti, poi rientrano nel torrente circolatorio e possono ritornare al midollo osseo o portarsi ai tessuti linfoidi periferici (alle tonsille, al tessuto linfoide dei tratti alimentari, respiratorio, alla milza, ai linfonodi). A questo punto le cellule si localizzano nei sinusoidi dei linfonodi e nella polpa bianca periarteriolare della milza; possono passare nel flusso linfatico generale, rientrare poi nel torrente circolatorio tramite il dotto toracico, il dotto linfatico di destra e le vene brachio cefaliche e, così, alla fine rientrare nuovamente ai tessuti linfatici. I linfociti B non passano dal timo, ma si portano direttamente, tramite la circolazione ai tessuti linfatici generali, dove si vanno a localizzare; nel loro percorso vanno incontro a fasi di differenziazione simile a quelle cui vanno incontro i linfociti T del timo, ma non è ancora ben chiaro dove queste fasi abbiano luogo.
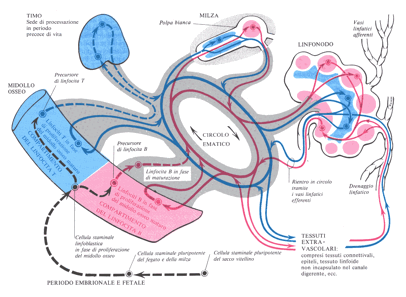
Schema che illustra gli attuali punti di vista sulle origini e sulla circolazione delle due maggiori classi di linfociti dal midollo osseo ai tessuti linfatici periferici; i linfociti B sono mostrati in rosso, i linfociti T in blu.









